Chromatin

| Übergeordnet |
|---|
| Chromosom |
| Untergeordnet |
| Euchromatin Heterochromatin zytoplasm./nukl. Chromatin aktives/ruhendes Chromatin |
| Gene Ontology |
| QuickGO |
Chromatin ist das Material, aus dem die Chromosomen bestehen. Es handelt sich um einen Komplex aus DNA und speziellen Proteinen, von denen wiederum etwa die Hälfte Histone sind. Der Name kommt von griech. chroma (Farbe), weil sich Chromatin mit basischen Kernfarbstoffen anfärben lässt. Im Lichtmikroskop erscheint es als sichtbares Fadengerüst im Zellkern einer eukaryotischen Zelle. Im funktionalen Sinn gilt alles, was sich während der Teilung des Zellkerns (Mitose oder Meiose) in den Chromosomen wiederfindet, als Chromatin – ausgenommen einige Strukturproteine. Chromatin ist neben den Nucleoli, der Kern-Grundsubstanz und der Kernhülle eine wichtige Strukturkomponente des Zellkerns (Nucleus).[1]
Chromatin besteht aus der DNA, die um die Histone gewickelt ist, sowie aus weiteren Proteinen, die sich an die DNA anlagern. DNA und Histone bilden die Nucleosomen, die kettenförmig aneinandergereiht sind. Die Nucleosomen werden mit Hilfe der Nichthiston-Proteine dichter gepackt. Chromatin ist somit das Produkt von Interaktionen der eukaryotischen DNA mit unterschiedlichen DNA-Bindeproteinen, die einen kompakten filamentösen Komplex bilden, den sogenannten Desoxyribonucleoprotein-Komplex, man spricht auch von Chromatinfasern oder Chromatinfäden (englisch: chromatin fibers). Durch die Komplexbildung werden die langen chromosomalen DNA-Stränge in ihrer Länge um das rund 10.000- bis 50.000-fache verkürzt (kondensiert), sodass sie in den Zellkern passen. Trotz der dichten Packung der DNA liegen die Chromosomen weiterhin in einer Form vor, die regulatorischen Proteinen Zugang zur DNA erlaubt, so dass die Biosynthese von RNA und Proteinen aus den genetischen Informationen (Genexpression) bzw. die Duplikation der chromosomalen DNA (Replikation) möglich ist.[2]
Während der Mitose und Meiose kondensieren die Chromosomen, so dass sie im Lichtmikroskop erkennbar werden. Die kleinsten lichtmikroskopisch sichtbaren Chromatinstrukturen nennt man Chromonema.
Das Verständnis der Chromatinstruktur und ihres Beitrags zu Regulation der Gene ist Gegenstand der Epigenetik.
Chromatinstrukturen machen Stäbchen bei nachtaktiven Säugetieren empfindlicher, da sie die Lichtausbreitung beeinflussen. Bei Nicht-Säugern ist das Phänomen noch nicht untersucht worden (Stand 2010).[3]
Chromatin-Typen
[Bearbeiten | Quelltext bearbeiten]Es werden zwei Typen von Chromatin unterschieden:
- Euchromatin, dessen DNA aktiv ist, d. h., zu Proteinen exprimiert werden kann. Die euchromatischen Abschnitte des Chromosoms weisen keine Unterschiede in ihrer Struktur auf, gleichgültig, in welchem Kondensationsgrad sich ein Chromosom befindet.
- Heterochromatin, das hauptsächlich aus inaktiver DNA besteht. Es scheint strukturelle Funktionen in den verschiedenen Kondensationsstufen auszuüben. Die heterochromatischen Abschnitte des Chromosoms weisen in der Interphase den gleichen Kondensationsgrad auf wie in der Metaphase, d. h., es bleibt auch im Interphasekern kondensiert und tritt in Form dichter Chromozentren in Erscheinung. Heterochromatin kann in zwei Untertypen unterteilt werden:
- Konstitutives Heterochromatin, das nie exprimiert wird. Es findet sich im Bereich des Centromers und besteht gewöhnlich aus repetitiven (sich wiederholenden) DNA-Sequenzen.
- Fakultatives Heterochromatin, das manchmal exprimiert wird.
Eine weitere begriffliche Abgrenzung kann somit auch nach den Kernteilungsphasen getroffen werden: Hierbei ist das Interphasechromatin gegenüber dem Metaphasechromatin mit seinen sehr kompakten Chromosomen stark aufgelockert.
Prokaryonten haben im Gegensatz zu Eukaryonten eine ringförmige DNA-Struktur. Die Eukaryonten haben Chromosomen, die die Struktur der DNA bilden.
Zeittafel wichtiger Entdeckungen
[Bearbeiten | Quelltext bearbeiten]
- 1842: Chromosom (Struktur) (Carl Wilhelm von Nägeli)
- 1874: Nukleinsäure (Friedrich Miescher)
- 1879: prägt Walther Flemming den Begriff Chromatin.
- 1883: August Weismann verbindet Chromatin mit der Vererbung.
- 1884: Albrecht Kossel entdeckt Histone.
- 1888: Sutton und Boveri schlagen die Theorie der Kontinuität von Chromatin während des Zellzyklus vor.[4]
- 1888: Wilhelm Waldeyer prägt den Begriff Chromosom
- 1910: Chromosomen sind die Träger der Gene (Thomas Hunt Morgan)
- 1928: Emil Heitz prägt den Begriff Heterochromatin und Euchromatin.
- 1942: Conrad Waddington postuliert die epigenetischen Landschaften.
- ca. 1945 Basenpaarung von Adenin und Thymin sowie Cytosin und Guanin postuliert (Erwin Chargaff, Chargaff-Regeln)
- 1948: Rollin Hotchkiss entdeckt DNA-Methylierung[5]
- 1953: Die DNA-Struktur wird aufgeklärt von James Watson, Francis Crick, Maurice Wilkins, Rosalind Franklin[6]
- 1961: Aufklärung der genetischen Struktur (Marshall Warren Nirenberg, Heinrich Matthaei)
- 1961: Mary Lyon postuliert das Prinzip der X-Inaktivierung.
- 1966: Histon-Modifikationen/-Acetylierung (Vincent Allfrey)
- 1973:/1974 Chromatinfasern werden entdeckt
- 1973:–75 Vom nu-Body zum Nukleosom (Ada Olins, Donald Olins, Roger Kornberg)
- 1975: Nukleosomen-Überstruktur/Solenoid (John T. Finch und Aaron Klug)
- 1975: Pierre Chambon prägt den Begriff der Nukleosomen.
- 1976: Chromatinfäden werden entdeckt[7]
- 1982: Chromosomenterritorien werden entdeckt[8]
- 1984: John T. Lis entwickelt die Chromatin-Immunpräzipitationstechnik.[9]
- 1996 wurde mit der Backhefe das erste Genom eines Eukaryonten veröffentlicht.[10]
- 2002: Job Dekker entwickelt die Chromosome Conformation Capture (3C) Technik.[11]
- 2006: Marieke Simons entwickelt die 4C-[12], Dostie die 5C-Methode[13]
- 2007: B. Franklin Pugh entwickelt die ChIP-Seq-Technik.[14]
- 2009: Lieberman-Aiden und Job Dekker erfindet die Hi-C-Technik[15], Melissa J. Fullwood erfindet die ChIA-Pet-Technik.[16]
- 2012: Eine Gruppe der Ren-Labs und die von Edith Heard und Job Dekker geleiteten Gruppen entdecken Topologically Associated Domains (TADs) bei Säugetieren.[17][18]
Siehe auch
[Bearbeiten | Quelltext bearbeiten]Literatur
[Bearbeiten | Quelltext bearbeiten]Evolution:
- R. Ammar, D. Torti u. a.: Chromatin is an ancient innovation conserved between Archaea and Eukarya. In: eLife. 1, 2012, S. e00078–e00078, doi:10.7554/eLife.00078.
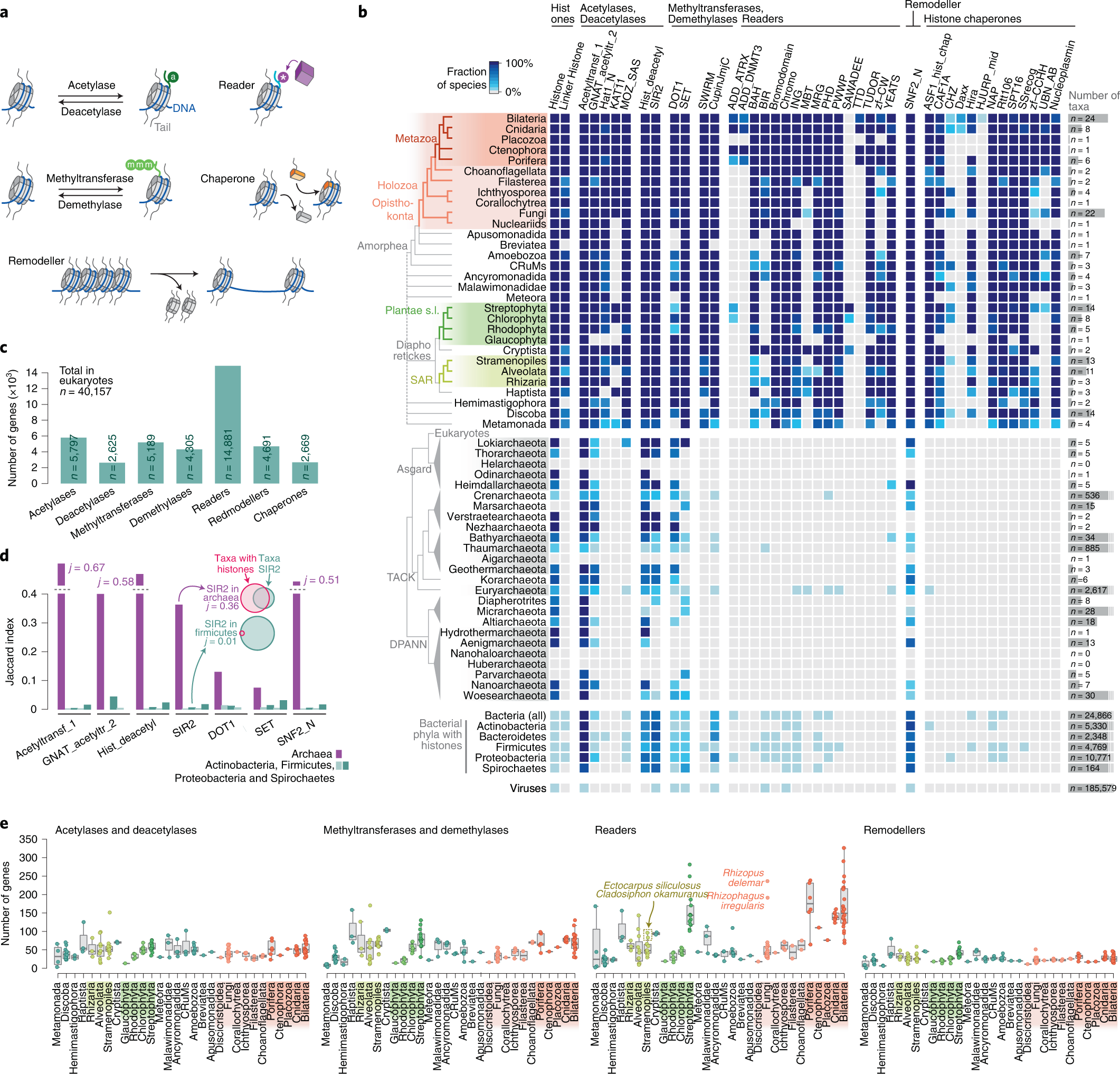
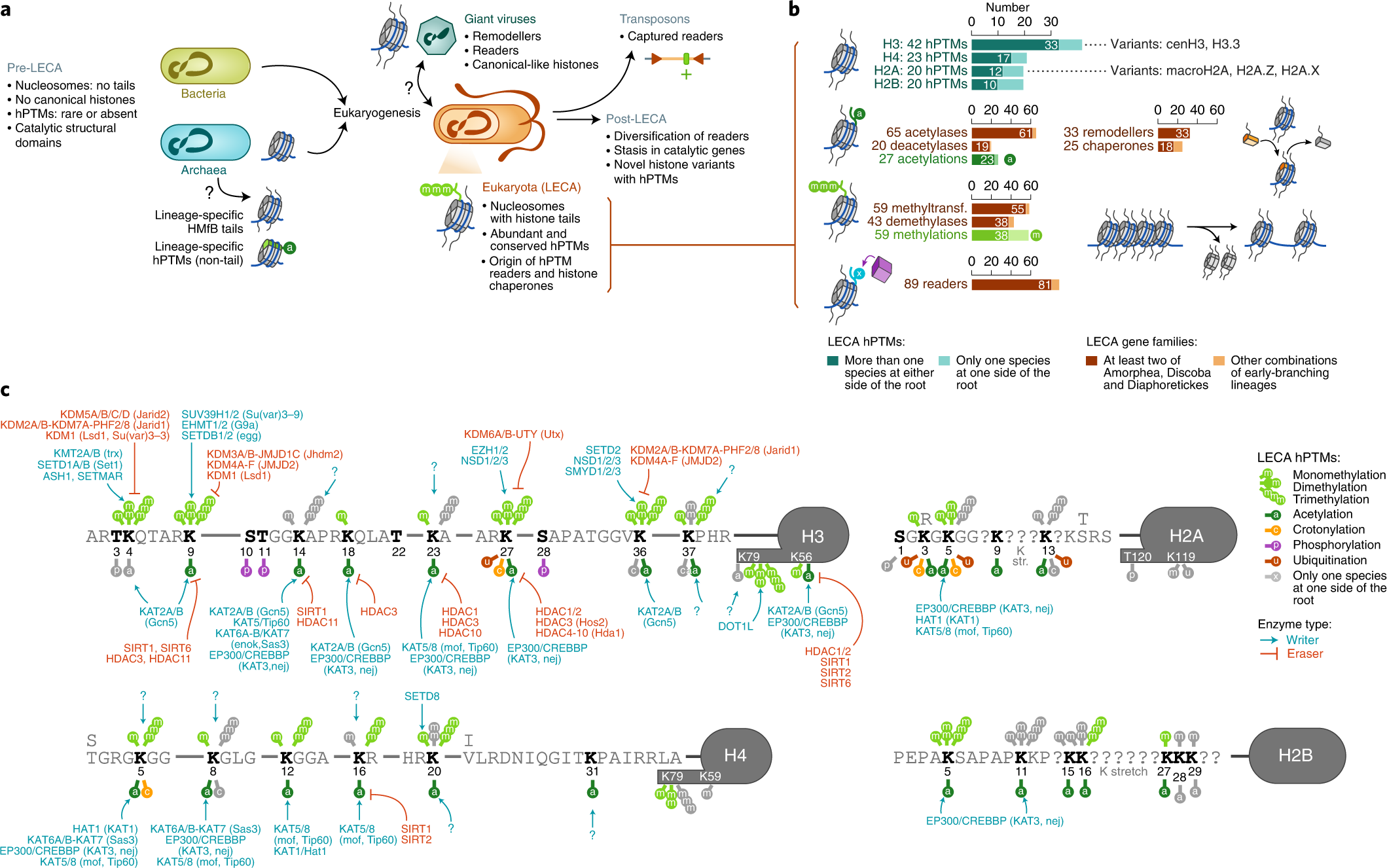
- Xavier Grau-Bové, Cristina Navarrete, Cristina Chiva, Thomas Pribasnig, Meritxell Antó, Guifré Torruella, Luis Javier Galindo, Bernd Franz Lang, David Moreira, Purificación López-Garcia, Iñaki Ruiz-Trillo, Christa Schleper, Eduard Sabidó, Arnau Sebé-Pedrós: A phylogenetic and proteomic reconstruction of eukaryotic chromatin evolution. In: Nature Ecology & Evolution, Band 6, S. 1007–1023, 9. Juni 2022; doi:10.1038/s41559-022-01771-6, PMID 35680998 PMC 7613034 (freier Volltext). Dazu:
- Fig. 1: Diversity of post-translational modifications in eukaryotic canonical and variant histones.
- Fig. 3: Taxonomic distribution of chromatin-associated gene classes.
- Fig. 6: Chromatin evolution and eukaryogenesis.
- Xavier Grau-Bové: Repository: Chromatin evolution. Auf: GitHub.
- Chromatin First Evolved in Ancient Microbes 1-2 Billion Years Ago, New Research Suggests. Auf: sci.news vom 13. Juni 2022.
- Shrouded in Mystery: Scientists Finally Discover the Origin of Chromatin. Auf: SciTechDaily vom 24. August 2022.
- Chromatin originated in ancient microbes one to two billion years ago. Genomic and proteomic analysis reveals that the regulatory role of chromatin is a eukaryotic innovation. Auf: ScienceDaily vom 9. Juni 2022. Quelle: Center for Genomic Regulation (CRG)
- Chromatin originated in ancient microbes one to two billion years ago. Center for Genomic Regulation (CRG) vom 9. Juni 2022.
{kind=link}
{kind=link}
{kind=link}
Histon-Modifikationen:
- V. G. Allfrey: Structural modifications of histones and their possible role in the regulation of ribonucleic acid synthesis. In: Proceedings. Canadian Cancer Conference. Band 6, 1966, S. 313–335, PMID 5934780.
- B. G. Pogo, A. O. Pogo, V. G. Allfrey, A. E. Mirsky: Changing patterns of histone acetylation and RNA synthesis in regeneration of the liver. In: Proceedings of the National Academy of Sciences of the United States of America. Band 59, Nr. 4, 1968, S. 1337–1344, PMC 224872 (freier Volltext).
Nukleosomen:
- A. L. Olins, D. E. Olins: Spheroid chromatin units (v bodies). In: Science. Band 183, Nr. 4122, 1974, S. 330–332, PMID 4128918.
Solenoid-Modell:
- J. T. Finch, A. Klug: Solenoidal model for superstructure in chromatin. In: Proceedings of the National Academy of Sciences of the United States of America. Band 73, Nr. 6, 1976, S. 1897–1901, PMC 430414 (freier Volltext).
Einzelnachweise
[Bearbeiten | Quelltext bearbeiten]- ↑ Hans Kleinig und Peter Sitte: Zellbiologie. Ein Lehrbuch. 3. Auflage, S. 176, Gustav Fischer Verlag (1992).
- ↑ The Chromatin Database: Chromatin and chromosome structure ( vom 22. Juli 2009 im Internet Archive) (Zugriff am 12. Juni 2009).
- ↑ schattenblick.de: Nachtsehen - Wenn jedes Lichtquant zählt. 17. April 2009.
- ↑ L.A.-C.P. Martins: Did Sutton and Boveri propose the so-called Sutton-Boveri chromosome hypothesis? In: Genetics and Molecular Biology. Band 22, Nr. 2, Juni 1999, ISSN 1415-4757, S. 261–272, doi:10.1590/S1415-47571999000200022 (scielo.br [abgerufen am 22. Juli 2019]).
- ↑ Haoyang Lu, Xinzhou Liu, Yulin Deng, Hong Qing: DNA methylation, a hand behind neurodegenerative diseases. In: Frontiers in Aging Neuroscience. Band 5, 2013, ISSN 1663-4365, doi:10.3389/fnagi.2013.00085, PMID 24367332, PMC 3851782 (freier Volltext).
- ↑ The Francis Crick Papers: The Discovery of the Double Helix, 1951-1953. Abgerufen am 22. Juli 2019.
- ↑ Ute Deichmann: Epigenetics: The origins and evolution of a fashionable topic. In: Developmental Biology. Band 416, Nr. 1, August 2016, S. 249–254, doi:10.1016/j.ydbio.2016.06.005 (elsevier.com [abgerufen am 22. Juli 2019]).
- ↑ T. Cremer, M. Cremer: Chromosome Territories. In: Cold Spring Harbor Perspectives in Biology. Band 2, Nr. 3, 1. März 2010, ISSN 1943-0264, S. a003889–a003889, doi:10.1101/cshperspect.a003889, PMID 20300217, PMC 2829961 (freier Volltext).
- ↑ D. S. Gilmour, J. T. Lis: Detecting protein-DNA interactions in vivo: distribution of RNA polymerase on specific bacterial genes. In: Proceedings of the National Academy of Sciences. Band 81, Nr. 14, 1. Juli 1984, ISSN 0027-8424, S. 4275–4279, doi:10.1073/pnas.81.14.4275, PMID 6379641, PMC 345570 (freier Volltext).
- ↑ Goffeau A, Barrell BG, Bussey H, R. W. Davis, B. Dujon, H. Feldmann, F. Galibert, J. D. Hoheisel, C. Jacq, M. Johnston, E. J. Louis, H. W. Mewes, Y. Murakami, P. Philippsen, H. Tettelin, S. G. Oliver: Life with 6000 genes. In: Science. 274. Jahrgang, Nr. 5287, Oktober 1996, S. 546, 563–7, doi:10.1126/science.274.5287.546, PMID 8849441, bibcode:1996Sci...274..546G (englisch).
- ↑ J. Dekker: Capturing Chromosome Conformation. In: Science. Band 295, Nr. 5558, 15. Februar 2002, S. 1306–1311, doi:10.1126/science.1067799.
- ↑ Marieke Simonis, Petra Klous, Erik Splinter, Yuri Moshkin, Rob Willemsen: Nuclear organization of active and inactive chromatin domains uncovered by chromosome conformation capture–on-chip (4C). In: Nature Genetics. Band 38, Nr. 11, November 2006, ISSN 1061-4036, S. 1348–1354, doi:10.1038/ng1896.
- ↑ J. Dostie, T. A. Richmond, R. A. Arnaout, R. R. Selzer, W. L. Lee: Chromosome Conformation Capture Carbon Copy (5C): A massively parallel solution for mapping interactions between genomic elements. In: Genome Research. Band 16, Nr. 10, 1. Oktober 2006, ISSN 1088-9051, S. 1299–1309, doi:10.1101/gr.5571506, PMID 16954542, PMC 1581439 (freier Volltext).
- ↑ Istvan Albert, Travis N. Mavrich, Lynn P. Tomsho, Ji Qi, Sara J. Zanton: Translational and rotational settings of H2A.Z nucleosomes across the Saccharomyces cerevisiae genome. In: Nature. Band 446, Nr. 7135, März 2007, ISSN 0028-0836, S. 572–576, doi:10.1038/nature05632.
- ↑ E. Lieberman-Aiden, N. L. van Berkum, L. Williams, M. Imakaev, T. Ragoczy: Comprehensive Mapping of Long-Range Interactions Reveals Folding Principles of the Human Genome. In: Science. Band 326, Nr. 5950, 9. Oktober 2009, ISSN 0036-8075, S. 289–293, doi:10.1126/science.1181369, PMID 19815776, PMC 2858594 (freier Volltext).
- ↑ Melissa J. Fullwood, Mei Hui Liu, You Fu Pan, Jun Liu, Han Xu: An oestrogen-receptor-α-bound human chromatin interactome. In: Nature. Band 462, Nr. 7269, November 2009, ISSN 0028-0836, S. 58–64, doi:10.1038/nature08497, PMID 19890323, PMC 2774924 (freier Volltext).
- ↑ Jesse R. Dixon, Siddarth Selvaraj, Feng Yue, Audrey Kim, Yan Li: Topological domains in mammalian genomes identified by analysis of chromatin interactions. In: Nature. Band 485, Nr. 7398, Mai 2012, ISSN 0028-0836, S. 376–380, doi:10.1038/nature11082, PMID 22495300, PMC 3356448 (freier Volltext).
- ↑ Elphège P. Nora, Bryan R. Lajoie, Edda G. Schulz, Luca Giorgetti, Ikuhiro Okamoto: Spatial partitioning of the regulatory landscape of the X-inactivation centre. In: Nature. Band 485, Nr. 7398, Mai 2012, ISSN 0028-0836, S. 381–385, doi:10.1038/nature11049, PMID 22495304, PMC 3555144 (freier Volltext).